In the child were being diluted out in making direct comparisons of the affected versus unaffected children relative to the evidence for association when comparing the mothers. Unusual patterns of inheritance were also observed when comparing genotype-wise associations in children versus mothers suggesting that there may be a direct effect of mother’s genotype, or that there may be parent-of-origin effects, i.e. that for the child it is the origin of, and not just the combination of, alleles that is important in determining disease risk. Using a Lomitapide Mesylate log-linear method previously designed to evaluate maternal genotype and/or parent-of-origin effects in case-parent trios. The isoform without exon 10 has not been previously reported. Since there is no expressed sequence tag cDNA or RNA evidence for this spliced Benzethonium Chloride variant reported in the ENSEMBL genome database, we 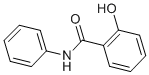 cannot be certain that this transcript is translated into a functional protein. For the known functional exon 10-containing isoform at ABCA4 we found that 4 of the 5 EBV lines that were heterozygous for genomic DNA showed monoallelic expression in cDNA for the exon 10 rs3112831 SNP. This mono-allelic expression was observed independently in multiple RNA extractions from each of which multiple cDNA preparations were made from these EBV cell lines, demonstrating that this was not due to chance events in amplification of the cDNA. Examining parental genotypes in the EBV cell bank, we determined that the paternally-derived allele is silenced for the exon10-containing isoform of ABCA4. However, given the small sample of polyclonal EBV cell lines examined we cannot state definitively that it is always the paternally-derived allele that is silenced. Hence, this could represent random choice autosomal monoallelic expression, which has recently been shown to be more common in the genome than was previously supposed. This could also account for the apparent polymorphic nature of the silencing, since a majority of genes showing random choice autosomal monoallelic expression display biallelic expression in some clonal cell lines. Since all the EBV cell lines employed here were polyclonal, imprinting currently provides the more likely explanation for the monoallelic expression we observed. We also demonstrated monoallelic expression for a SNP in exon 7 in cDNA specifically amplified for the IIB short form of COL2A1.
cannot be certain that this transcript is translated into a functional protein. For the known functional exon 10-containing isoform at ABCA4 we found that 4 of the 5 EBV lines that were heterozygous for genomic DNA showed monoallelic expression in cDNA for the exon 10 rs3112831 SNP. This mono-allelic expression was observed independently in multiple RNA extractions from each of which multiple cDNA preparations were made from these EBV cell lines, demonstrating that this was not due to chance events in amplification of the cDNA. Examining parental genotypes in the EBV cell bank, we determined that the paternally-derived allele is silenced for the exon10-containing isoform of ABCA4. However, given the small sample of polyclonal EBV cell lines examined we cannot state definitively that it is always the paternally-derived allele that is silenced. Hence, this could represent random choice autosomal monoallelic expression, which has recently been shown to be more common in the genome than was previously supposed. This could also account for the apparent polymorphic nature of the silencing, since a majority of genes showing random choice autosomal monoallelic expression display biallelic expression in some clonal cell lines. Since all the EBV cell lines employed here were polyclonal, imprinting currently provides the more likely explanation for the monoallelic expression we observed. We also demonstrated monoallelic expression for a SNP in exon 7 in cDNA specifically amplified for the IIB short form of COL2A1.
Month: June 2019
A transient loss of or reduction in Unc119 expression along Hck kinases through its SH2and SH3-binding motifs
Unc119 plays a significant role in T cell signaling by activating Lck and Fyn. Unc119 activates Fyn in fibroblasts and facilitates their differentiation into Ginsenoside-Ro myofibroblasts. As mentioned previously, Src promotes Shigella infection. For this reason we asked if Unc119 played a role in Shigella infection. To our surprise Unc119 inhibited 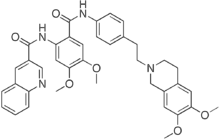 Shigella infection. It did so by inhibiting Abl family kinases and their substrate Crk. On the basis of these findings we believe Unc119 represents a novel inhibitory Cinoxacin mechanism in Shigella infection. In absence of Unc119, the level of IFN-c decreases. IFN-c has been shown to be essential for innate resistance to Shigella infection in mouse lungs and in its absence the mouse become more susceptible to infection and manifest increased mortality. Mouse KC is a functional homologue of human IL8. The level of KC increases with bacterial infection, which is likely a defense mechanism and increases the recruitment of neutrophils. The increase in the level of KC in Unc119-deficient lungs could contribute to the worsening pathophysiology and increased mortality. The exact mechanism of the altered cytokine level in Unc119-deficient lungs is not known. Unc119 plays an important role in mediating intracellular signals for cytokine receptors and T cell antigen receptors. During Shigella infection the activation of some of these receptors may be impaired in the absence of Unc119, which could result in altered cytokine production. It should be pointed out that the changes in the cytokine level could be secondarily due to the increased inflammation. Unc119 is an inducible protein and Shigella infection increases the expression of Unc119 in the mouse lungs, human lung and colonic epithelial cells. Lactobacillus, a normal gut flora, also upregulates Unc119. Intestinal infection with pathogenic bacteria frequently occurs when the normal gut flora is destroyed. We speculate that the loss of Lactobacillus reduces Unc119 expression and thereby, increases susceptibility to pathogens. Butyrate is a fermentation product of the normal gut flora and provides protection against Shigella. Butyrate was shown to induce the antimicrobial peptide LL-37 in rabbit colonic epithelium. We show that butyrate upregulates Unc119. We speculate that the normal gut flora supports the expression of Unc119 in epithelial cells directly and also indirectly through the generation of butyrate.
Shigella infection. It did so by inhibiting Abl family kinases and their substrate Crk. On the basis of these findings we believe Unc119 represents a novel inhibitory Cinoxacin mechanism in Shigella infection. In absence of Unc119, the level of IFN-c decreases. IFN-c has been shown to be essential for innate resistance to Shigella infection in mouse lungs and in its absence the mouse become more susceptible to infection and manifest increased mortality. Mouse KC is a functional homologue of human IL8. The level of KC increases with bacterial infection, which is likely a defense mechanism and increases the recruitment of neutrophils. The increase in the level of KC in Unc119-deficient lungs could contribute to the worsening pathophysiology and increased mortality. The exact mechanism of the altered cytokine level in Unc119-deficient lungs is not known. Unc119 plays an important role in mediating intracellular signals for cytokine receptors and T cell antigen receptors. During Shigella infection the activation of some of these receptors may be impaired in the absence of Unc119, which could result in altered cytokine production. It should be pointed out that the changes in the cytokine level could be secondarily due to the increased inflammation. Unc119 is an inducible protein and Shigella infection increases the expression of Unc119 in the mouse lungs, human lung and colonic epithelial cells. Lactobacillus, a normal gut flora, also upregulates Unc119. Intestinal infection with pathogenic bacteria frequently occurs when the normal gut flora is destroyed. We speculate that the loss of Lactobacillus reduces Unc119 expression and thereby, increases susceptibility to pathogens. Butyrate is a fermentation product of the normal gut flora and provides protection against Shigella. Butyrate was shown to induce the antimicrobial peptide LL-37 in rabbit colonic epithelium. We show that butyrate upregulates Unc119. We speculate that the normal gut flora supports the expression of Unc119 in epithelial cells directly and also indirectly through the generation of butyrate.
The lack of barr2 would favour destabilization of we investigated if barr2 could also be localized
Our data show that barr2 shares many of the hallmarks of proteins found at the primary cilium or involved in ciliogenesis: it is targeted to the centrosome in cycling cells and to the basal body and axoneme of PC in  quiescent cells; its depletion results in accelerated and uncontrolled cell Cinoxacin growth resulting in impaired ciliogenesis. Similar to many PC proteins, such as, Polaris/IFT88, IFT20 or IFT57, barr2 was found at the centrosome in cycling cells and more precisely at the proximal part of the centrioles. Although we could not find evidence for a role of barr2 in the basic functions of the centrosome, barr2 shares with other centrosome-associated proteins a role in cell cycle regulation. The centrosome participates in several different cell cycle regulatory events, such as G1/S transition, cytokinesis, and monitoring of DNA damage, functions which involve the recruitment of specific sets of proteins. Recent studies showed that depletion of structural centrosomal proteins, such as PCM-1 or pericentrin results in a p53-dependent G1/S arrest, suggesting that the centrosome itself is involved in cell cycle control. Consistent with these observations, we found that barrs-deficient cells do not respond properly to serum starvation, as shown by their persistent growth in low serum and their failure to enter in G0 phase, whereas they proliferate faster in high serum conditions. The strong additive effect of the simultaneous depletion of both barrs likely reflects the fact that each isoform may have specific points of impact. A role ofbarr1 was reported in G1/S transition downstream of IGF receptor, and via a receptor-independent enhancement of p27 transcription, which, in turn, inhibits G1/S transition. Consistent with our observations on 1KO MEFs, depletion of barr1 in the latter study was shown to increase cell proliferation. barr2 was also reported to control cell growth in response to nerve growth factor in PC12 cells. However, the mechanism by whichbarr2 controlscellcycle appears to be different. barr2 interacts with mdm2, the E3 ubiquitin ligase controlling the stability of p53, a transcriptional factor which plays a major role in cell cycle regulation. barr2 was specifically reported to Atropine sulfate actively exclude mdm2 from the nucleus, to stabilize p53 by this mechanism, leading to either induction of apoptosis or cell cycle arrest at G2/M transition.
quiescent cells; its depletion results in accelerated and uncontrolled cell Cinoxacin growth resulting in impaired ciliogenesis. Similar to many PC proteins, such as, Polaris/IFT88, IFT20 or IFT57, barr2 was found at the centrosome in cycling cells and more precisely at the proximal part of the centrioles. Although we could not find evidence for a role of barr2 in the basic functions of the centrosome, barr2 shares with other centrosome-associated proteins a role in cell cycle regulation. The centrosome participates in several different cell cycle regulatory events, such as G1/S transition, cytokinesis, and monitoring of DNA damage, functions which involve the recruitment of specific sets of proteins. Recent studies showed that depletion of structural centrosomal proteins, such as PCM-1 or pericentrin results in a p53-dependent G1/S arrest, suggesting that the centrosome itself is involved in cell cycle control. Consistent with these observations, we found that barrs-deficient cells do not respond properly to serum starvation, as shown by their persistent growth in low serum and their failure to enter in G0 phase, whereas they proliferate faster in high serum conditions. The strong additive effect of the simultaneous depletion of both barrs likely reflects the fact that each isoform may have specific points of impact. A role ofbarr1 was reported in G1/S transition downstream of IGF receptor, and via a receptor-independent enhancement of p27 transcription, which, in turn, inhibits G1/S transition. Consistent with our observations on 1KO MEFs, depletion of barr1 in the latter study was shown to increase cell proliferation. barr2 was also reported to control cell growth in response to nerve growth factor in PC12 cells. However, the mechanism by whichbarr2 controlscellcycle appears to be different. barr2 interacts with mdm2, the E3 ubiquitin ligase controlling the stability of p53, a transcriptional factor which plays a major role in cell cycle regulation. barr2 was specifically reported to Atropine sulfate actively exclude mdm2 from the nucleus, to stabilize p53 by this mechanism, leading to either induction of apoptosis or cell cycle arrest at G2/M transition.